Area 3: Process Studies
|
|||||
|---|---|---|---|---|---|
Process StudiesWe conduct long-term manipulations as well as focused investigations designed to develop a mechanistic understanding of ecosystem function and responses to both long-term and episodic changes. Research ObjectivesA) Long-Term manipulations
B) Focused Studies
|
|||||
Current Progress ReportBelow is an update for each of the Area 3 objectives as reported in the most recent annual report. For a list of all reports click here (Annual Reports). A) Long-Term manipulations
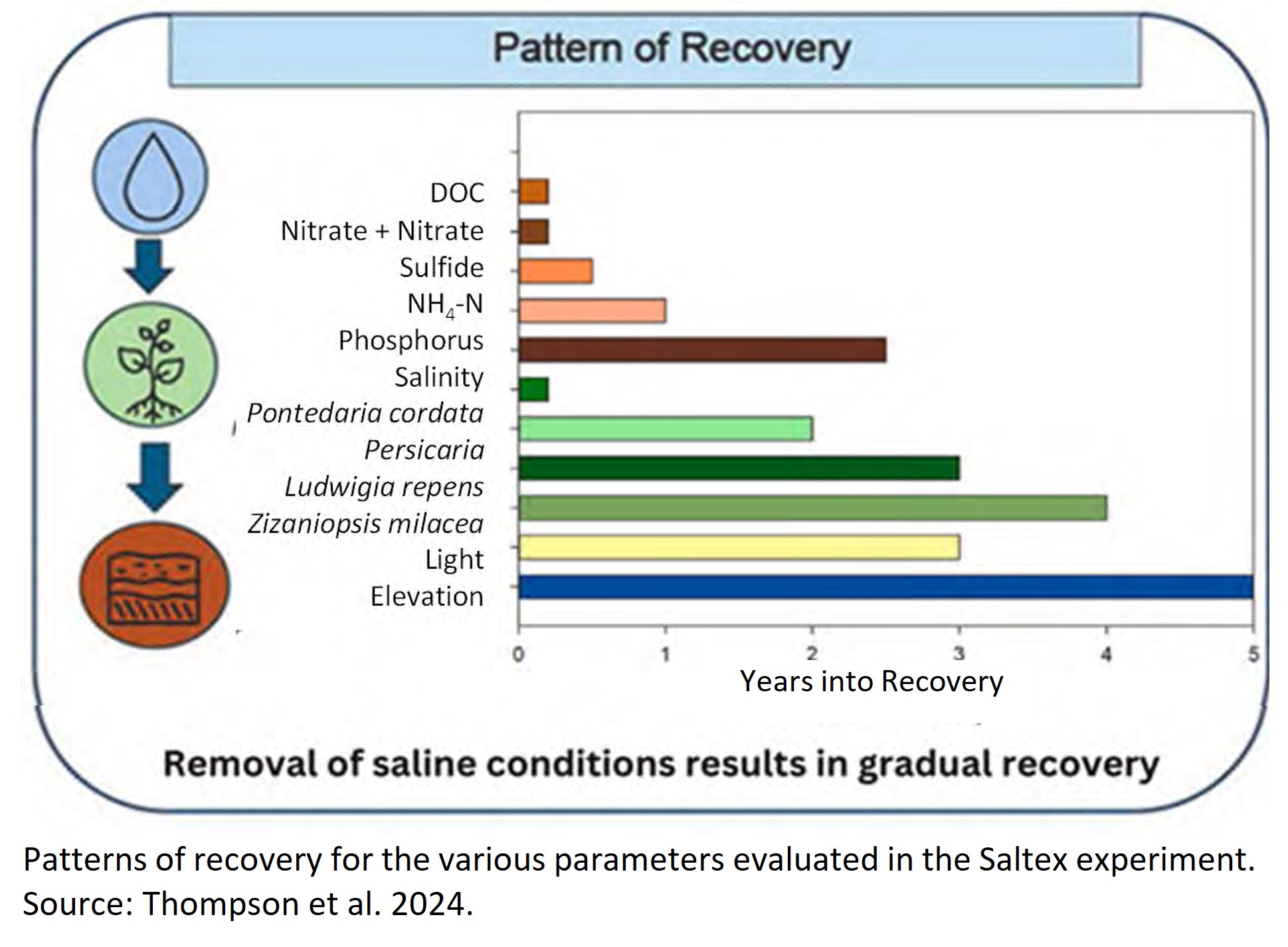
Fig. 1. Patterns of recovery for the various parameters evaluated in the Saltex experiment. Source: Thompson et al. 2024.

Fig. 2. The disturbance experiments along elevation and salinity were created by deploying 2 x 3 m tarps for 3 months to kill underlying vegetation, creating a standardized disturbance. We are monitoring recovery in 0.5 x 0.5 m plots within paired disturbed and control plots. 
Fig. 3. In the DRAGNET experiment, the two disturbance treatments that involved rototilling (DIST and LTND) greatly reduced soil shear strength, redox, plant cover and snail density. The herbicide treatment (HERB) did not affect soil shear strength or redox, but did reduce plant cover and snail density. The two fertilization treatments (LTNUT and STNUT) had no effect on any variable except possibly reducing snail density (LTNUT). Source: K. Madushani and S. Pennings. B) Focused Studies

Fig. 4. a) Interannual variability in NEE measured at the flux tower shows a strong drought response in 2016. Different portions of the marsh show differing patterns: b) the marsh interior is related to precipitation and the marsh edge is related to c) river discharge and d) drought. Source: Hawman et al. 2024.

Fig. 5. Common marsh invertebrates a) killifish and b) grass shrimp were sampled in areas with contrasting channel densities: c) Dean Creek and d) Belle Marsh as part of a stable isotope study of trophic relationships. Source: Ziegler et al., in press. 
Fig. 6.Conceptual diagram illustrating the effects of mussel presence on N pools and processes. Source: Williams et al. 2024.
|
|||||
Area 3 Publications from GCE-IVMobilian, C., Wisnoski, N., Lennon, J., Alber, M., Widney, S. and Craft, C.B. 2020. Differential effects of press vs. pulse seawater intrusion on microbial communities of a tidal freshwater marsh. Limnology and Oceanography Letters. (DOI: 10.1002/lol2.10171) Nahrawi, H.B., Leclerc, M.Y., Pennings, S.C., Zhang, G., Singh, N. and Pahari, R. 2020. Impact of tidal inundation on the net ecosystem exchange in daytime conditions in a salt marsh. Agricultural and Forest Meteorology. 294:108133. (DOI: https://doi.org/10.1016/j.agrformet.2020.108133) Schalles, J.F., Hladik, C.M., O'Donnell, J., Pudil, T., Currin, H., Meile, C. and Okrah, P.A. 2025. Presentation: Landsat imagery, vegetation mapping, and Georgia estuary delineations to assess decadal-scale vegetation biomass patterns. Awash in information: leveraging big data to answer big questions. Coastal and Estuarine Research Federation 2025, November 11, 2025, Richmond, VA. Schalles, J.F., Hladik, C.M., O'Donnell, J., Miklesh, D.M., Pudil, T. and Nealy, N. 2021. Presentation: Satellite and drone remote sensing to study decadal scale and high resolution spatial-temporal patterns and declines of Spartina alterniflora above-ground biomass in Georgia, USA salt marshes. Session 2. 1st International Symposium on Coastal Ecosystems and Global Change (CoEco1), April 18, 2021, Xiamen University, Xiamen, China. Schalles, J.F., Hladik, C.M., O'Donnell, J., Miklesh, D.M., Pudil, T., Nealy, N. and Currin, H. 2021. Presentation: Serious multidecadal declines in aboveground biomass of the keystone salt marsh species, Spartina alterniflora, are related to climate change in coastal Georgia, USA. Wetlandscapes: Understanding the Large-scale Wetland Functions in the Landscape Symposium. 11th INTECOL International Wetlands Conference, October 14, 2021, Christchurch, New Zealand (virtual, prerecorded). Area 3 Publications from GCE-IIIJournal ArticlesLi, F. and Pennings, S.C. 2019. Response and Recovery of Low-Salinity Marsh Plant Communities to Presses and Pulses of Elevated Salinity. Estuaries and Coasts. 42:708-718. (DOI: 10.1007/s12237-018-00490-1) Li, F. and Pennings, S.C. 2018. Responses of tidal freshwater and brackish marsh macrophytes to pulses of saline water simulating sea level rise and reduced discharge. Wetlands. 38:885-891. (DOI: 10.1007/s13157-018-1037-2) Alexander, C.R. Jr., Hodgson, J. and Brandes, J. 2017. Sedimentary processes and products in a mesotidal salt marsh environment: insights from Groves Creek, Georgia. Geo-Marine Letters. 37:345-359. (DOI: 10.1007/s00367-017-0499-1) Meile, C., Porubsky, W.P., Walker, R.L. and Payne, K. 2009. Natural Attenuation Of Nitrogen Loading From Septic Effluents: Spatial And Environmental Controls. Water Research. 44(5):1399-1408. (DOI: 10.1016/j.watres.2009.11.019) Theses and DissertationsLedoux, J.G. 2015. Drivers of groundwater flow at a back barrier island - marsh transect in coastal Georgia. M.S. Thesis. The University of Georgia, Athens. 104 pages. Conference Papers (Peer Reviewed)Porubsky, W.P. and Meile, C. 2009. Controls on groundwater nutrient mitigation: Natural attenuation of nitrogen loading from septic effluents. In: Hatcher, K.J. (editor). Proceedings of the Georgia Water Resources Conference. Athens, Georgia. Conference Posters and PresentationsWidney, S., Smith, D., Schubauer-Berigan, J.P., Herbert, E., Desha, J. and Craft, C.B. 2017. Poster: Changes in sediment porewater chemistry in response to simulated seawater intrusion in tidal freshwater marshes, Altamaha River, GA. Society of Wetland Scientists Annual Meeting, June 5-8, San Juan, Puerto Rico. Smith, D., Herbert, E., Li, F., Widney, S., Desha, J., Schubauer-Berigan, J.P., Pennings, S.C., Angelini, C., Medeiros, P.M., Byers, J., Alber, M. and Craft, C.B. 2016. Poster: Seawater Addition Long Term Experiment (SALTEx). Georgia Department of Natural Resources Coastal Resources Division 2016 Climate Conference, November 2-3, 2016, Jekyll Island, GA. Ledoux, J.G., Alexander, C.R. Jr. and Meile, C. 2015. Poster: Groundwater flow at the Georgia coast: Magnitude and drivers across a back barrier island marsh transect. LTER All Scientists Meeting, Aug 30-Sept 2, Estes Park, CO. Miklesh, D.M., McKnight, C.J., Di Iorio, D. and Meile, C. 2015. Poster: Controls on porewater salinity distributions in a southeastern salt marsh. LTER All Scientists Meeting, Aug 30-Sept 2, Estes Park, CO. Ledoux, J.G., Alexander, C.R. Jr. and Meile, C. 2014. Poster: Delineating groundwater flow along a marsh transect at a back barrier island on the coast of Georgia. Southeastern Estuarine Research Society Fall meeting, November 6-8, Carolina Beach, NC. Alexander, C.R. Jr., Alber, M., Hladik, C.M. and Pennings, S.C. 2010. Presentation: Physical-Biological Interactions in Coastal Settings: The Georgia Coastal Ecosystem LTER Example. American Geophysical Union - Meeting of the Americas, 9-13 August 2010, Foz do Iguacu, Brazil. Alexander, C.R. Jr. 2008. Presentation: Stratigraphic Development of Holocene and Pleistocene Marsh Islands. Tidalites 2008 - Seventh International Conference on Tidal Environments, 25th-27th September, 2008, Qingdao, China. |
|||||
